一、遗传变异与饮食文化差异
嗅觉受体是人类对食品和外界环境香气感知主要接收器,在人类进化和环境选择过程中嗅觉基因的遗传变异对香气认知和灵敏度有重要影响。嗅觉受体的基因序列多样性已被证明有助于食物偏好,Eriksson等通过对14604名欧洲人全基因组关联分析发现,位于11号染色体嗅觉受体基因簇rs72921001存在对醛类香气敏感的嗅觉受体OR6A2基因,导致对香菜排斥。Gisladottir等证实TAAR5基因变异可影响冰岛人对鱼腥味的认知和感知,部分个体携带的特定TAAR5基因突变可使其感知的鱼腥味减弱,甚至觉得鱼腥味令人愉悦。此外,OR6C70基因变体可使个体对甘草香气的感知能力更强烈,并对其感知更愉悦,这种变异在东亚比在欧洲更常见。
饮食文化与遗传变异存在必然联系,饮食文化决定了香气感知的偏好,而偏好会促进遗传变异,进而反馈调节香气感知。不同国家和地区的饮食文化存在显著的多元性和差异性,如西方人认为豆类食品应该划分为咸味食品,然而在亚洲地区豆制品虽然可作为咸味食品,但在多数情况下被划分为甜味食品。Gotow等证实文化差异会驱动消费者增强对熟悉食品的后鼻腔香气感知,同时也伴随着食品味觉感知的强化。
饮食文化与遗传变异存在必然联系,饮食文化决定了香气感知的偏好,而偏好会促进遗传变异,进而反馈调节香气感知。不同国家和地区的饮食文化存在显著的多元性和差异性,如西方人认为豆类食品应该划分为咸味食品,然而在亚洲地区豆制品虽然可作为咸味食品,但在多数情况下被划分为甜味食品。Gotow等证实文化差异会驱动消费者增强对熟悉食品的后鼻腔香气感知,同时也伴随着食品味觉感知的强化。
二 、香气传递路线差异
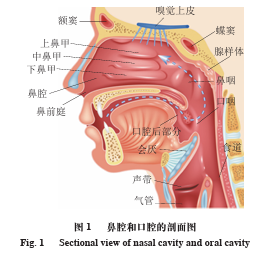
鼻腔香气感知通过鼻孔将外界环境中的香气分子吸入鼻腔,传输至嗅觉上皮后与嗅觉受体结合,从而形成前鼻腔香气感知。后鼻腔香气感知主要源于食品口腔加工过程中释放的香气,通过呼吸气流将口腔中的香气分子传输至口腔后部,再经口咽部和鼻咽传输到后鼻腔区域,与嗅觉上皮的嗅觉受体结合,最终形成后鼻腔香气感知(图1)。前鼻香气与外部环境有关,后鼻腔香气伴随着口腔加工行为,体现了口腔的内部状态。